重组蛋白表达中包涵体的形成与优化策略
重组蛋白广泛应用于基础研究、生物制药和工业酶制剂等领域。大肠杆菌(Escherichia coli)因生长快、操作简便、成本低、表达量高,长期作为主要的重组蛋白宿主。然而高水平表达时,目的蛋白常因折叠失败而形成不可溶聚集体——包涵体,导致活性丧失,需经复杂的溶解/复性才能恢复,显著降低回收率与经济性。尽管包涵体常被视为“副产物”,近年来对其结构、半活性包涵体以及工艺优化的研究表明,在特定情况下,包涵体生产-复性已成为可行策略。
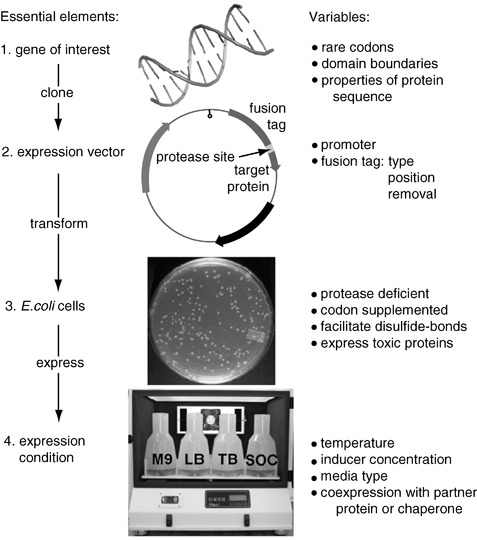
蛋白包涵体的特征
包涵体是细胞内由目标蛋白(或部分非折叠、错误折叠蛋白)聚集成的密集颗粒结构,通常为球形或椭球形,直径在 0.2–1.5 µm 之间,在电子显微镜下表现为高密度颗粒。它们在常规裂解缓冲条件下表现为不溶性,在离心后多出现在沉淀中。尽管传统观点认为包涵体中蛋白已被彻底变性,但越来越多研究表明,包涵体内部可能保留部分二级结构或折叠中间态(native‐like 或半折叠结构),某些包涵体甚至具有一定的残余活性。这种含有部分可折叠结构或松散聚集态的包涵体,有时称作“非经典包涵体”(non-classical IBs)。
包涵体具有以下典型特性:
* 高度富集目标重组蛋白,杂质较少;
* 较高的稳定性和抗降解能力;
* 颗粒密度较高,便于通过离心等手段回收;
* 溶解和复性难度较大,易在复性过程中再聚集。
这些特性使得包涵体既是挑战,也可能在一定条件下成为优势(特别是在高表达、低杂质回收等方向)。
包涵体的形成机制与影响因素
包涵体的形成本质上是蛋白错误折叠与聚集,即细胞“蛋白稳态”(proteostasis) 失衡的结果。在高表达压力下,伴侣和折叠酶系统饱和、翻译过快、中间态不稳、分子挤密、疏水作用及二硫键错误配对等共同作用,使部分蛋白偏向聚集。
主要影响因素包括:
1. 表达水平(拷贝数、启动子、诱导浓度):速率过高会超出折叠负荷,增加错误折叠。
2. 诱导与培养条件(温度、时间、浓度):高温、快速或强诱导促进包涵体;低温、缓诱导则减少聚集。
3. 宿主与伴侣系统:不同菌株(如 BL21、C41/C43 等)差异明显;伴侣蛋白(DnaK-DnaJ, GroEL-GroES, TF, ClpB 等)可辅助折叠,降低聚集。
4. 密码子与翻译速率:非最优密码子致速率不均,易聚集;优化密码子或补充稀有 tRNA 可改善。
5. 氨基酸序列特性:疏水残基、低复杂区、β-聚集域、多半胱氨酸等均增加聚集风险。
6. 胞内环境(pH、盐度、氧化还原状态、挤密效应等):如高盐、极端 pH 或还原环境都会影响折叠稳定性。
包涵体优化策略
- 上游表达阶段优化
1.1 降低表达速率(“慢表达”策略):通过降低温度(如 20–30 °C)、减少 IPTG 浓度(0.1–0.5 mM)及合理选择诱导时间,可减缓翻译速度,给肽链更多折叠时间,减少聚集。Box–Behnken 优化设计已证实该策略可提升可溶比例。
1.2 调控启动子与拷贝数:使用弱启动子(lac、tac、弱型 T7)或低拷贝/复制控制型质粒,可避免折叠体系超载,降低包涵体形成。
1.3 自诱导与流加培养:自诱导体系通过乳糖/葡萄糖控制在高密度时自动表达,速率温和;流加培养缓慢供给诱导剂,避免强诱导引发聚集。
1.4 共表达伴侣与折叠助剂:共表达 DnaK-DnaJ、GroEL-GroES、TF、ClpB 等可增强折叠能力,减少聚集。培养基中添加甘氨酸、甜菜碱、山梨醇、乙醇、金属离子或辅因子也能改善折叠。
1.5 宿主株选择/工程菌株:优化宿主株如 C41/C43、BL21-pLysS、BL21Star、ArcticExpress 等,可提高可溶表达。工程化改造折叠网络或削弱应激通路也能增强折叠效率。
1.6 蛋白工程策略:① 融合标签:如 MBP、GST、SUMO、Trx 提高可溶性,降低聚集。② 序列改造:去除疏水尾段、低复杂区或聚集片段;定点突变降低聚集风险。③ 融合诱导包涵体:在需要高产纯化时,可用特定标签(如 PagP)刻意诱导包涵体形成。
- 包涵体性质调控与“软化”策略
对于那些仍然以包涵体形式表达、难以获得高比例可溶性的蛋白,其次的思路是通过“性质调控”使包涵体更易被溶解/复性。主要方法包括:
2.1 控制表达温度与诱导条件以获得松散结构包涵体
低温诱导、低诱导强度使得包涵体结构相对松软,内部仍保留部分可折叠单元,更容易溶解。
2.2 促进非经典包涵体的生成
通过温和诱导条件(如较低表达速率、添加辅助分子、使用温和去折叠剂)使包涵体生成状态更接近中间状态,从而更易通过溶剂提取活动性蛋白,而非完全变性重构。
2.3 共表达“解聚因子”或蛋白质量控制因子
如共表达 ClpB 等解聚伴侣,可能在包涵体形成期间干预聚集路径,从而降低聚集密度或形成较松散颗粒。
2.4 利用辅助小分子、添加剂调节包涵体结构
在表达培养过程中向培养基中添加低浓度的弱变性剂(如低浓度尿素、糖类、甘油、入渗性保护剂等),有时可以使蛋白在聚集时处于较松散状态。
- 包涵体溶解与复性策略
当包涵体形成后,关键在于如何高效溶解并正确复性获得活性蛋白。传统方法多用高浓度变性剂(如 6–8 M 尿素、6 M 盐酸胍)彻底解聚,但往往破坏已有二级结构,复性过程中易再聚集,回收率低。近年来发展出温和溶解策略,利用低浓度变性剂、非变性洗涤剂、极端 pH 或压力处理等方式,尽量减少结构破坏,从而提高复性效率。
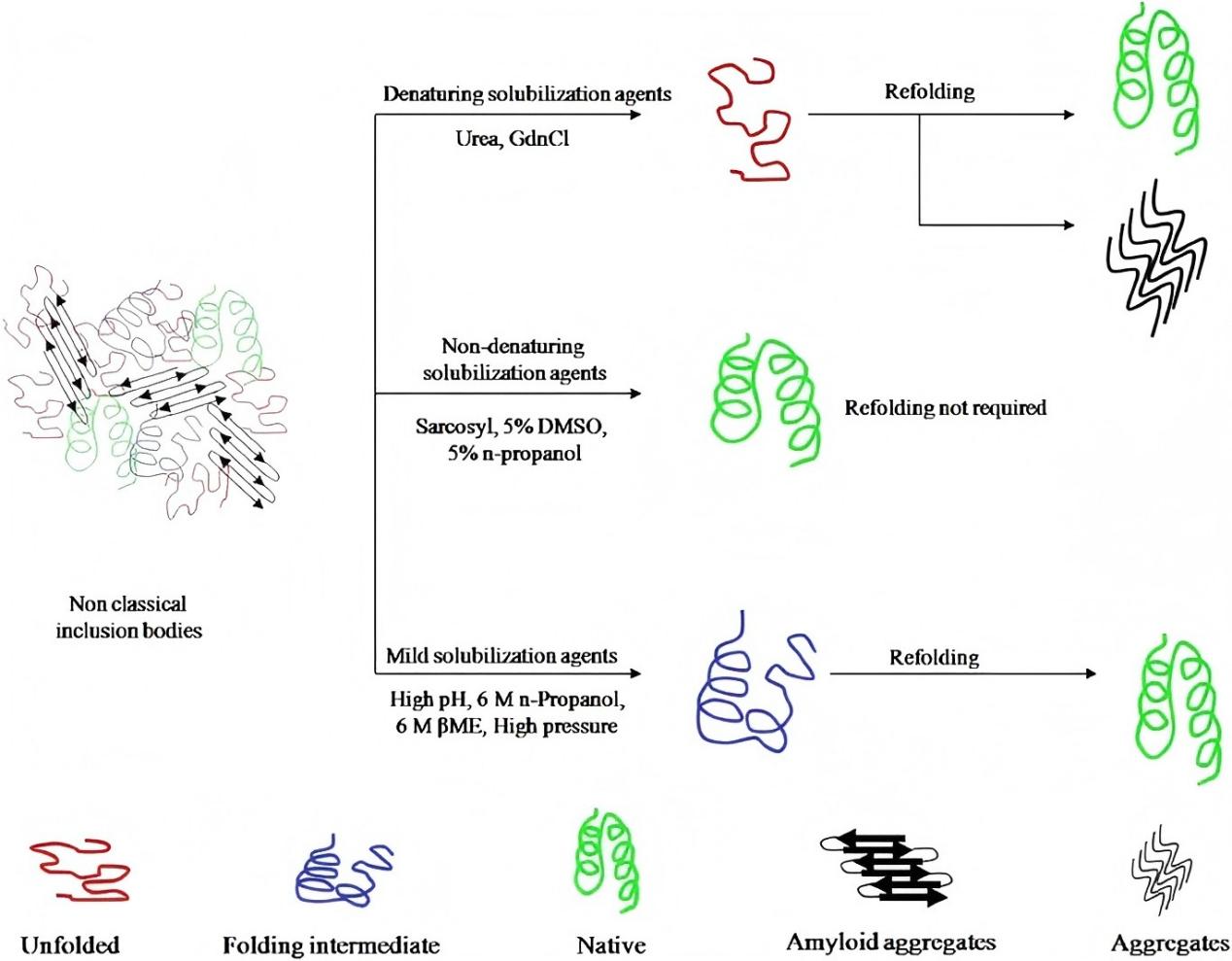
主要方法包括:
3.1 低浓度变性剂 + 助剂
例如使用 2–4 M 尿素联合去污剂(如 N-lauroyl sarcosine)、甜菜碱、非离子洗涤剂、甘油等,使包涵体部分解聚,同时尽可能保留二级结构。
3.2 碱性高 pH 溶解
利用高 pH(如 > 12)条件,可能在弱变性剂辅助下促使聚集体松散解离,已被报道可用于某些蛋白。
3.3 高压辅助溶解
施加高静水压(如 2–4 kbar)可破坏非共价聚集键,配合温和条件下复性,有助于恢复折叠。
3.4 有机溶剂辅助
在低浓度有机溶剂(如 5% n-丙醇、DMSO 等)辅助下结合弱变性剂实现包涵体部分溶解。此类方法在一些研究中表现出较高复性率。
3.5 缓慢稀释或梯度稀释复性
将溶解态蛋白缓慢稀释至低变性剂浓度、逐步移除助剂,以降低聚集风险。
3.6 辅助分子辅助折叠
在复性缓冲中添加分子伴侣(如 GroEL/ES、DnaK 系统)、折叠助剂(如血红素、金属离子、氧化还原对、促进形成二硫键的复性体系)等,以增强复性效率。
3.7 优化温度、离子强度与缓冲体系
选择适合的温度、pH、盐浓度等条件,以平衡折叠速率与抑制聚集。通常较低温度(如 4–15°C)、适中盐浓度、缓冲稳定性较好的离子环境较有利。
- 下游纯化与整合流程优化
在包涵体溶解与复性后,尚需进一步纯化以去除宿主蛋白、折叠错误聚集体等。优化策略包括:
中间分级纯化:利用分子排阻、亲和层析、离子交换等手段,在复性过程中早期去除杂质或错误折叠体;
在线复性-纯化整合:将复性与层析结合设计(如折叠-色谱)、使用梯度洗脱减少再聚集;
微流控与连续化技术:如近年报道的微流控复性装置,可提高折叠效率、降低聚集;Frontiers 的文章指出采用两段串联反应器进行连续包涵体表达/回收可提升产率与时空产能。
自动化高通量筛选:建立高通量评分体系(如浊度法)筛选最优复性条件。
应用实例与案例回顾
1. 酶类蛋白(工业酶)
某些真菌来源的木聚糖酶在 E. coli 中高水平表达时主要形成包涵体。研究者通过降低诱导温度(37 °C → 20 °C)、减少 IPTG 浓度,并在复性过程中添加甘油和谷胱甘肽对/氧化还原体系,提高了可溶性复性蛋白比例,恢复了较高的酶活。
2. 含二硫键的蛋白(如抗体片段、干扰素)
哺乳动物来源的干扰素在原核系统中因二硫键形成困难,大多沉淀为包涵体。实验中,研究者采用包涵体提取—尿素溶解—稀释复性的方法,并在复性缓冲液中加入氧化谷胱甘肽和还原谷胱甘肽配比体系,促进正确二硫键形成,最终获得有活性的干扰素。
3. 膜蛋白(转运蛋白)
膜蛋白在 E. coli 中常以包涵体形式沉积。某实验通过在培养基中添加渗透保护剂(山梨醇、甘氨酸甜菜碱),并共表达分子伴侣 GroEL/ES,使部分膜蛋白折叠得以改善;其余形成的包涵体则通过温和溶解剂(N-月桂酰肌氨酸钠)溶解后复性,获得部分功能性蛋白。
4. 病毒蛋白(神经氨酸酶)
病毒来源的神经氨酸酶在 E. coli 中主要以包涵体形式表达。通过优化诱导时间点、IPTG 浓度和表达周期,获得较高表达量。进一步结合温和溶解与复性流程,成功恢复了酶学活性,为疫苗研究提供了实验基础。
5. 人源蛋白(生长因子)
人源表皮生长因子在大肠杆菌中过量表达后几乎全部沉淀为包涵体。研究者利用 SUMO 融合标签改善部分可溶性,同时对包涵体部分进行低浓度尿素溶解,并在复性缓冲液中添加甘油和精氨酸抑制再聚集,最终获得高纯度的活性蛋白。
在重组蛋白表达过程中,包涵体确实是一个需要面对但并非不可逾越的障碍。通过对包涵体形成机制的深入理解,并结合表达调控、宿主设计、融合工程、温和溶解-复性策略及下游纯化流程优化,研究者能够获得较为理想的功能蛋白产出。未来,随着自动化、高通量、计算辅助方法的发展,包涵体优化工艺有望更加高效、可推广,将在生物制剂和工业酶制备中发挥更大作用。